Deux processus neuroinflammatoires concomitants sont à l'origine de l'accumulation du handicap dans la SEP1-5
De plus en plus de preuves suggèrent que les neuroinflammations aiguë et chronique sont à l'origine de l'accumulation du handicap dès le début de la maladie, avec une contribution sous-estimée de cette dernière1-5
Regardez comment la progression apparaît
Découvrez comment les neuroinflammations aiguë et chronique à bas bruit contribuent à l'accumulation du handicap.


La neuroinflammation aiguë, provoquée en partie par les lymphocytes B et T activés provenant de la périphérie, entraîne des poussées, des lésions aiguës et parfois un handicap résiduel lié aux poussées (RAW).1,3

La neuroinflammation chronique à bas bruit, liée en partie à la présence de cellules microgliales à l’état pathologique dans le SNC, se manifeste cliniquement par une progression indépendante des poussées (PIRA).1,3
La physiopathologie de la neuroinflammation aiguë et chronique6,7
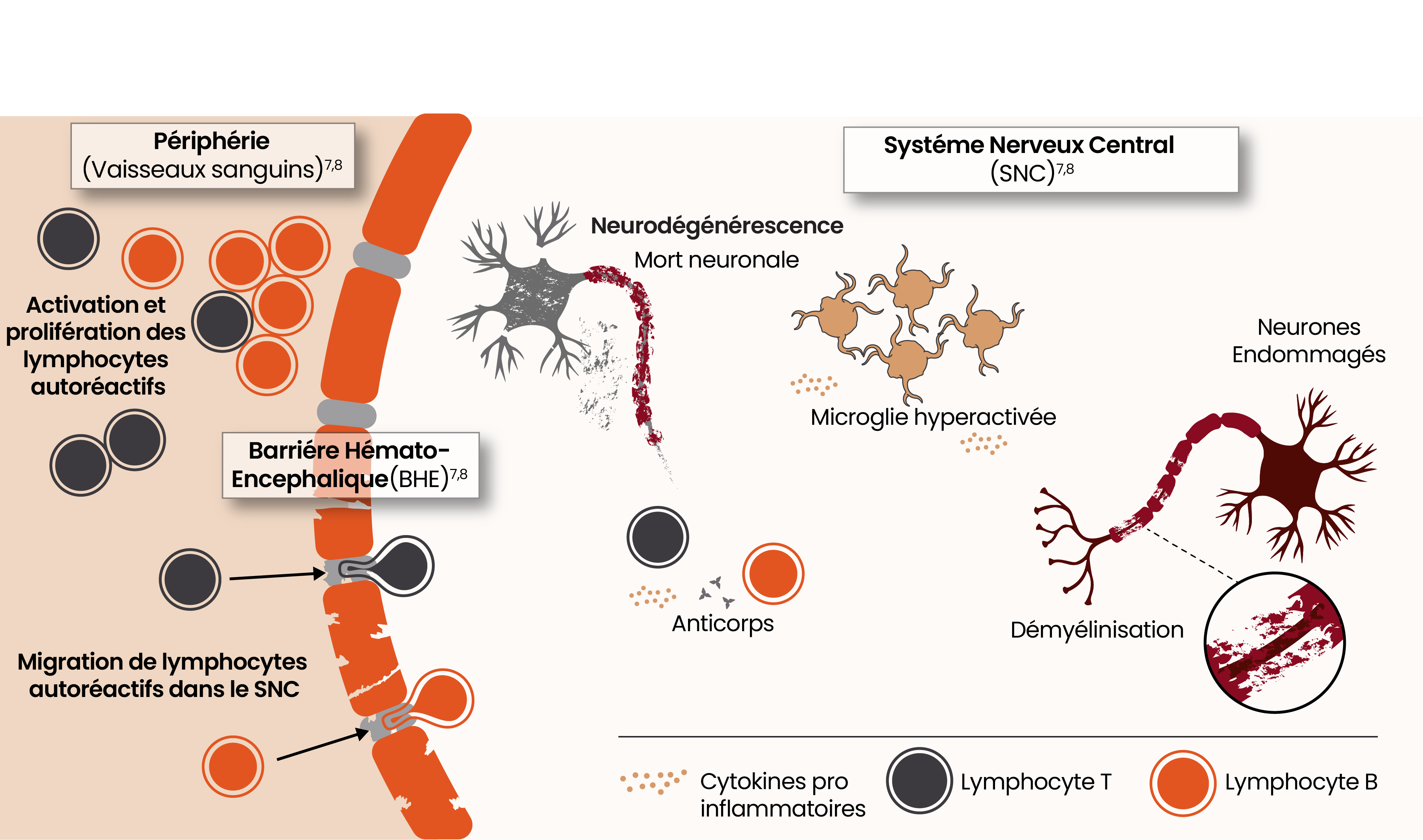
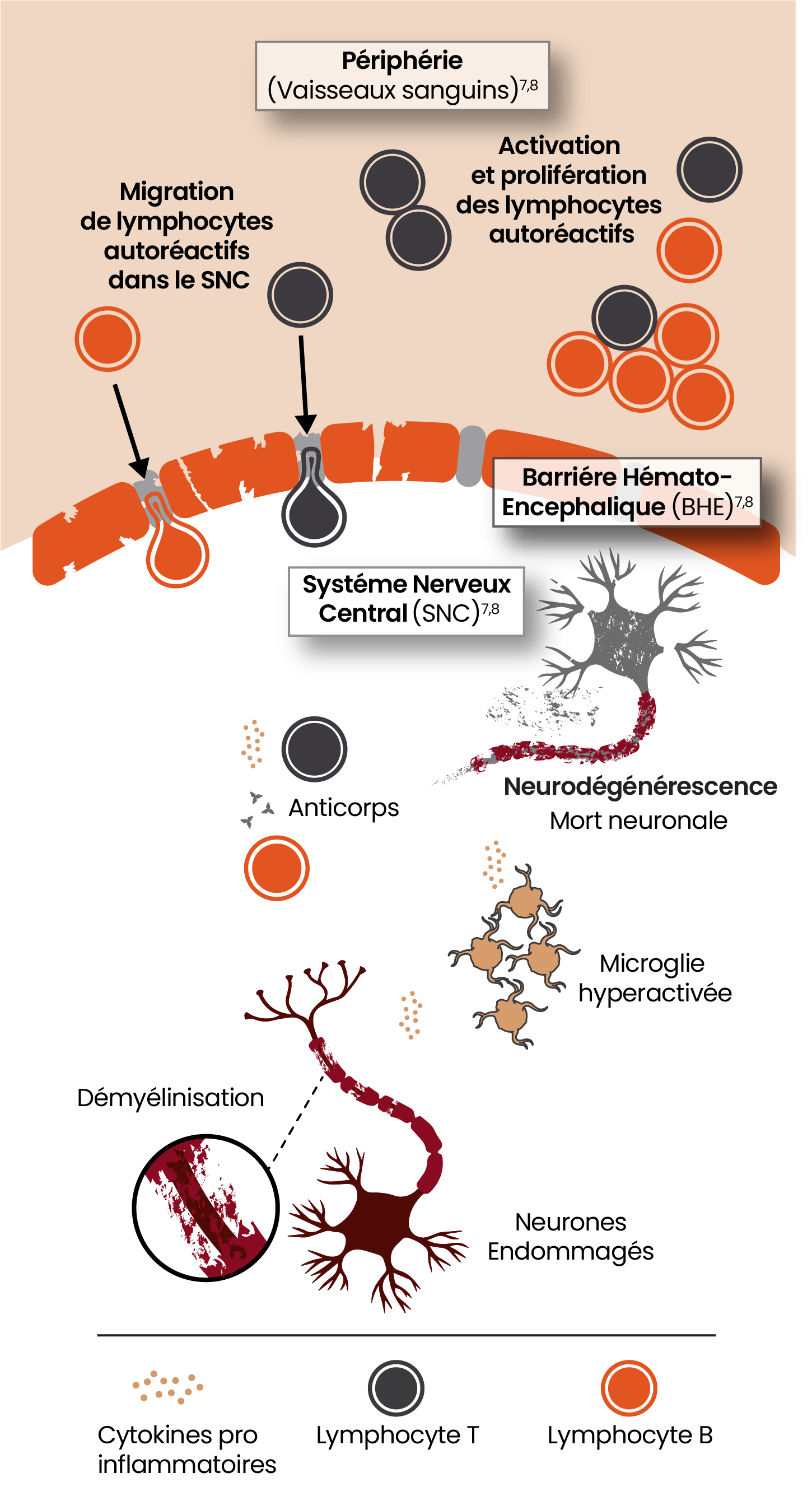
Historiquement, la SEP était considérée comme une maladie médiée par les cellules de l’immunité adaptative, provenant de la périphérie, telles que les lymphocytes B et T. L’évolution de notre compréhension porte sur le rôle essentiel joué par les cellules de l’immunité innée dans le SNC, telles que les cellules microgliales, sur l'accumulation du handicap.8
L'activité des cellules microgliales, dans le SNC, joue un rôle fondamental dans la smoldering MS, à l’origine d’une accumulation du handicap1,8-11

La microglie est hyperactivée dans la SEP et jouerait un rôle important dans l'accumulation du handicap.12,13

Les cellules microgliales chargées en fer se situent au niveau de la bordure des lésions paramagnétiques* (PRL – Paramagnetic rim lesions) et sont associées à une augmentation du handicap dans les SEP-RR et SP.14
La perte synaptique induite par la microglie hyperactivée a été associée à une perte cognitive.15-17
*Les PRLs sont un type de lésions chroniques actives
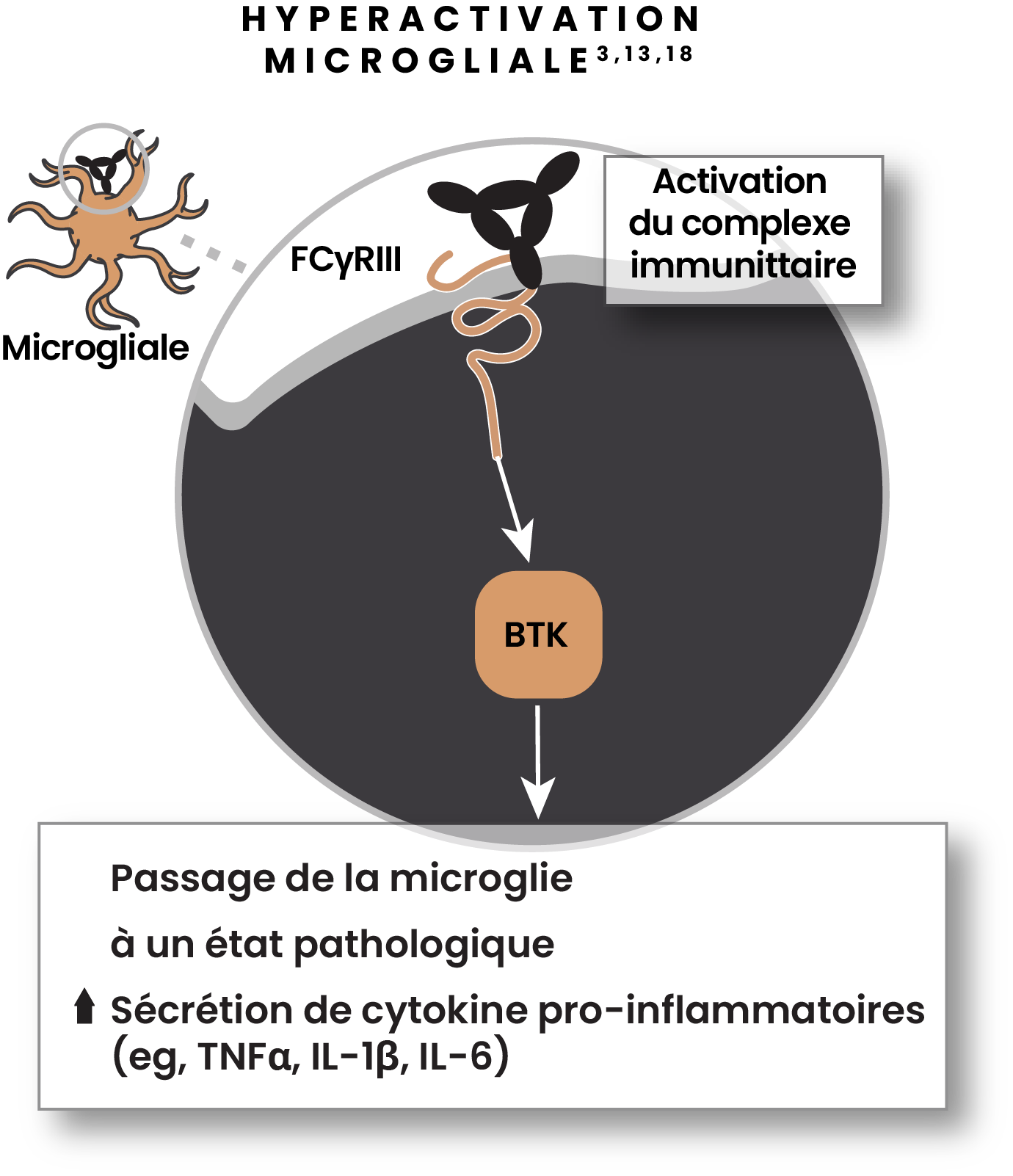
L'enzyme Tyrosine Kinase de Bruton (BTK) est une nouvelle voie de recherche3,13,18
La BTK joue un rôle primordial dans l'activation des lymphocytes B et de la microglie dans la SEP3,13,18
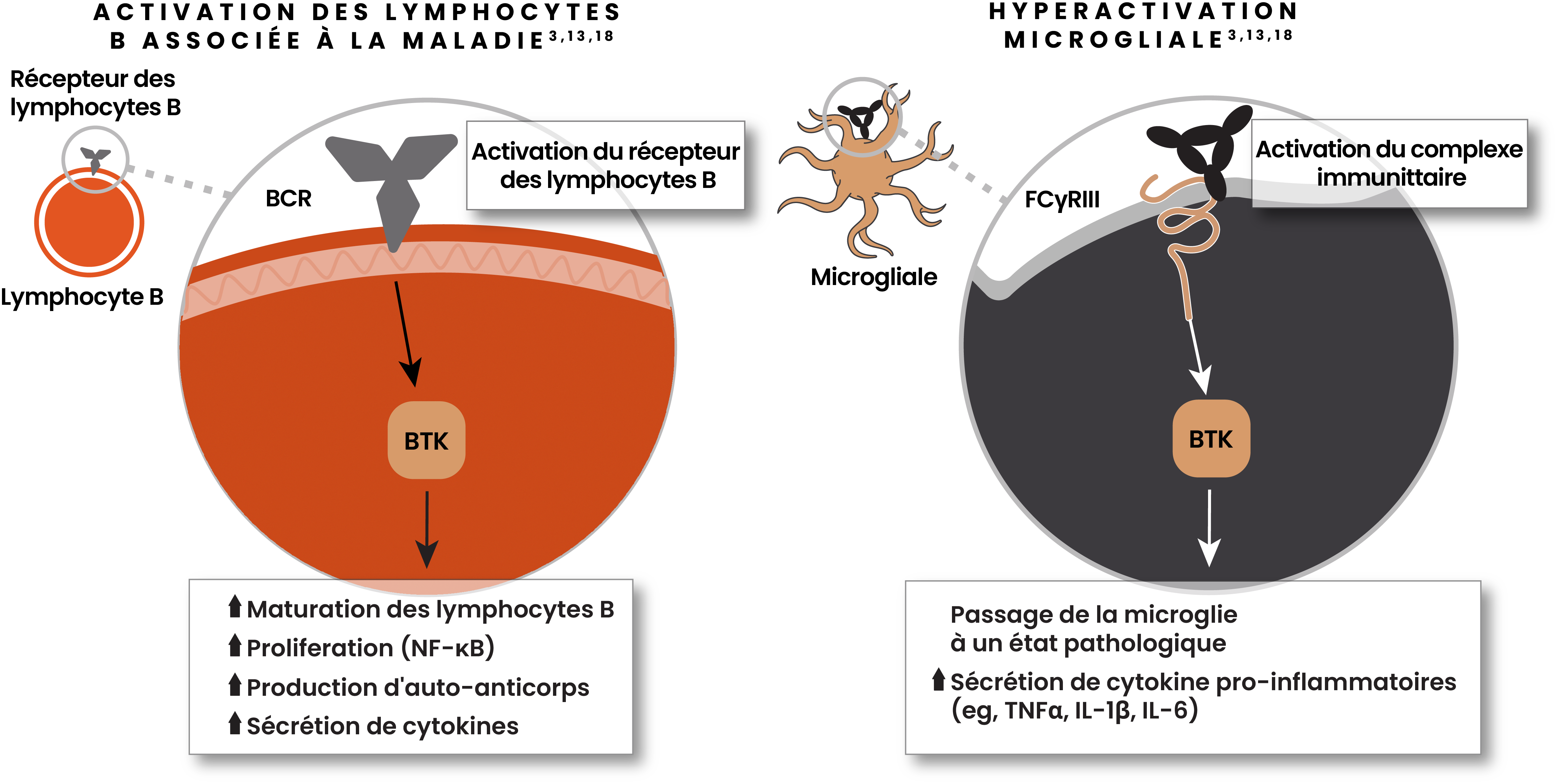
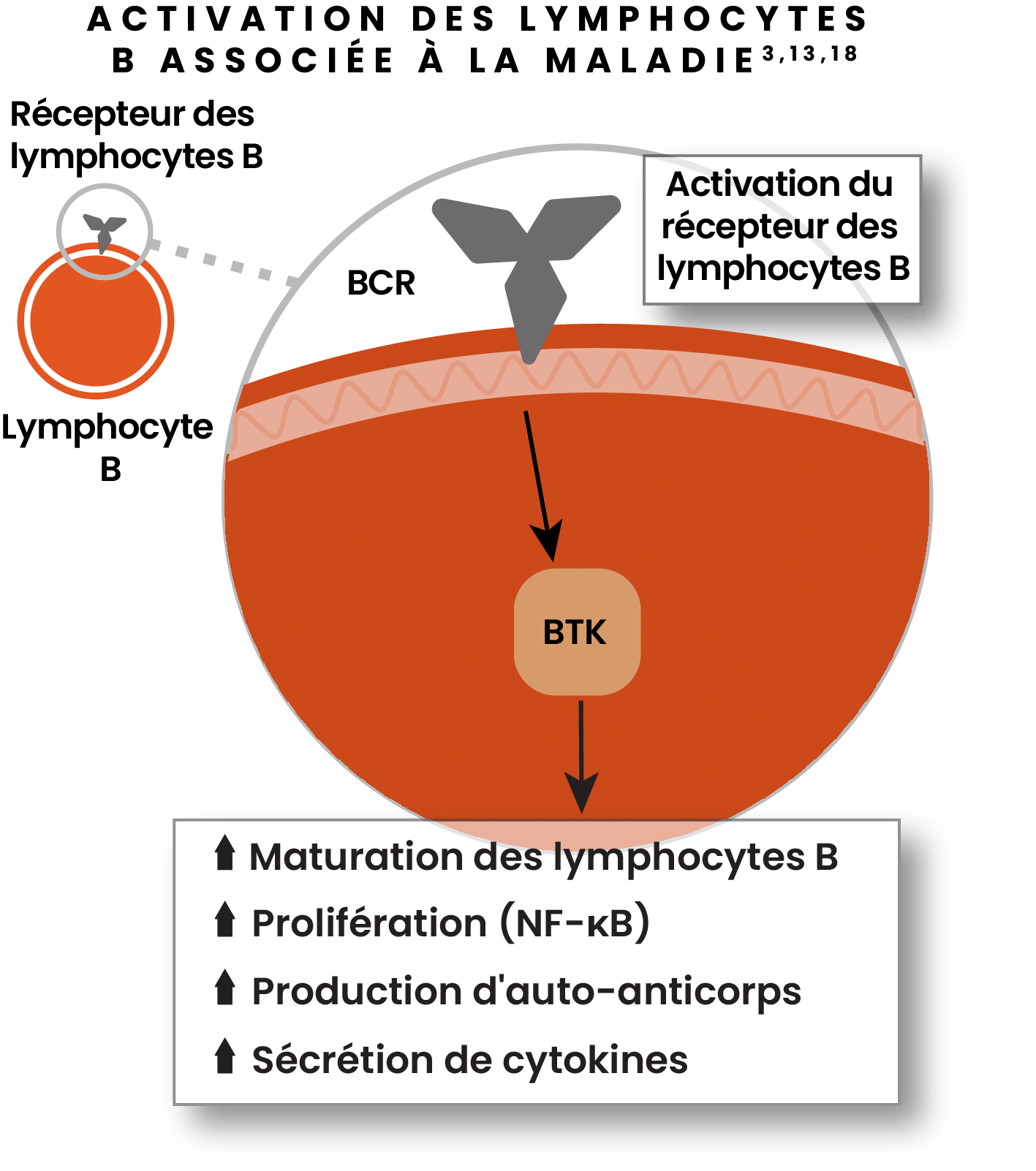
- Dans les lymphocytes B, la BTK favorise la prolifération, la production d'anticorps et la sécrétion de cytokines3,13,18
- L'enzyme Tyrosine Kinase de Bruton (BTK) est une nouvelle voie de recherche3,13,18
- La BTK est fortement exprimée dans la microglie au sein des tissus lésionnels des patients atteints de SEP-SP3,13,18
Dès les premiers stades précoces de la SEP, la microglie peut passer d’un état homéostatique à un état pathogénique20
Les cellules microgliales, dans leur état pathogénique, sont associées à une perte axonale, à une neurodégénérescence, à une perte de volume cérébral et à l'accumulation du handicap à long terme.21,22
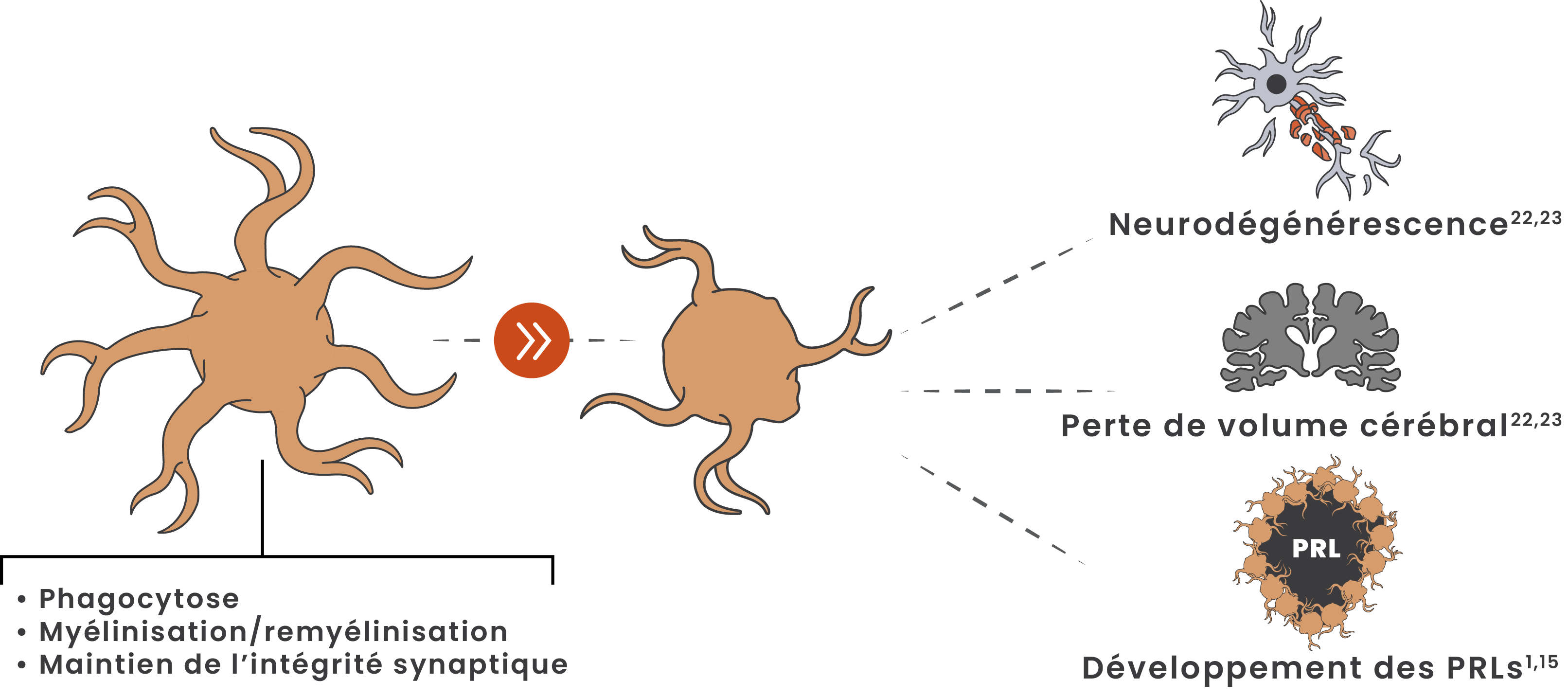
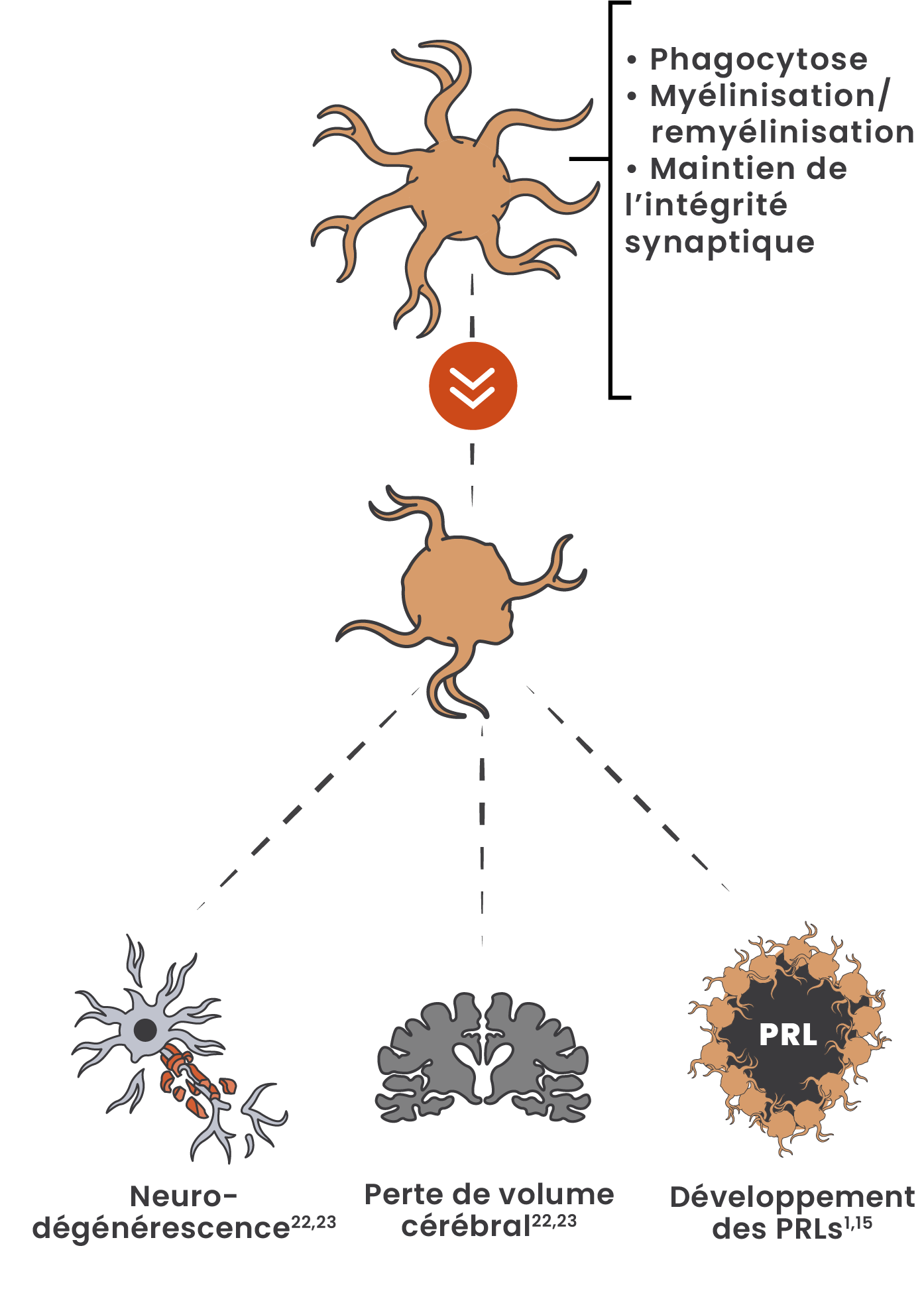
Adapté de Geladaris et al. 2021

Continuez d’explorer
Comprendre le rôle de chaque processus
Références:
-
Giovannoni G, Popescu V, Wuerfel J, et al. Smouldering multiple sclerosis: the ‘real MS’. Ther Adv Neurol Disord. 2022;15:17562864211066751. doi:10.1177/17562864211066751
-
Giovannoni G. The neurodegenerative prodrome in multiple sclerosis. Lancet Neurol. 2017;16(6):413-414.
-
Häusser-Kinzel S, Weber MS. The role of B cells and antibodies in multiple sclerosis, neuromyelitis optica, and related disorders. Front Immunol. 2019;10:201. doi:10.3389/fimmu.2019.00201
-
Krieger SC, Antoine A, Sumowski JF. EDSS 0 is not normal: multiple sclerosis disease burden below the clinical threshold. Mult Scler. 2022;28(14):2299-2303. doi:10.1177/13524585221108297
-
Filippi M, Amato MP, Centonze D, et al. Early use of high-efficacy disease-modifying therapies makes the difference in people with multiple sclerosis: an expert opinion. J Neurol. 2022;269(10):5382-5394.
-
Gandhi R, Laroni A, Weiner HL. Role of the innate immune system in the pathogenesis of multiple sclerosis. J Neuroimmunol. 2010;221(1-2):7-14.
-
Reich DS, Lucchinetti CF, Calabresi PA. Multiple sclerosis. N Engl J Med. 2018;378(2):169-180.
-
Hernández-Pedro NY, Espinosa-Ramirez G, de la Cruz VP, Pineda B, Sotelo J. Initial immunopathogenesis of multiple sclerosis: innate immune response. Clin Dev Immunol. 2013;2013:413-465. doi:10.1155/2013/413465
-
Pilz G, Sakic I, Wipfler P, et al. Chemokine CXCL13 in serum, CSF and blood-CSF barrier function: evidence of compartment restriction. Fluids Barriers CNS. 2020;17(1):7. doi:10.1186/s12987-020-0170-5
-
Matejuk A, Ransohoff RM. Crosstalk between astrocytes and microglia: an overview. Front Immunol. 2020;11:1416. doi:10.3389/fimmu.2020.01416
-
Margoni M, Preziosa P, Filippi M, Rocca MA. Anti-CD20 therapies for multiple sclerosis: current status and future perspectives. J Neurol. 2022;269(3):1316-1334.
-
Kamma E, Lasisi W, Libner C, Ng HS, Plemel JR. Central nervous system macrophages in progressive multiple sclerosis: relationship to neurodegeneration and therapeutics. J Neuroinflammation. 2022;19(1):45. doi:10.1186/s12974-022-02408-y
-
Gruber, R.C., Wirak, G.S., Blazier, A.S. et al. BTK regulates microglial function and neuroinflammation in human stem cell models and mouse models of multiple sclerosis. Nat Commun 15, 10116 (2024)
-
Absinta M, Sati P, Masuzzo F, et al. Association of chronic active multiple sclerosis lesions with disability in vivo. JAMA Neurol. 2019;76(12):1474-1483.
-
Friese MA. Widespread synaptic loss in multiple sclerosis. Brain. 2016;139(pt 1):2-4.
-
Jürgens T, Jafari M, Kreutzfeldt M, et al. Reconstruction of single cortical projection neurons reveals primary spine loss in multiple sclerosis. Brain. 2016;139(pt 1):39-46.
-
Werneburg S, Jung J, Kunjamma RB, et al. Targeted complement inhibition at synapses prevents microglial synaptic engulfment and synapse loss in demyelinating disease. Immunity. 2020;52(1):167-182.
-
Keaney J, Gasser J, Gillet G, Scholz D, Kadiu I. Inhibition of Bruton's Tyrosine Kinase modulates microglial phagocytosis: therapeutic implications for Alzheimer's disease. J Neuroimmune Pharmacol. 2019;14(3):448-461.
-
Hendriks RW. Drug discovery: new BTK inhibitor holds promise. Nat Chem Biol. 2011;7(1):4-5.
-
Guerrero BL, Sicotte NL. Microglia in multiple sclerosis: friend or foe? Front Immunol. 2020;11:374. doi:10.3389/fimmu.2020.00374
-
Datta G, Colasanti A, Rabiner EA, et al. Neuroinflammation and its relationship to changes in brain volume and white matter lesions in multiple sclerosis. Brain. 2017;140(11):2927-2938. doi:10.1093/brain/awx228
-
Geladaris A, Häusler D, Weber MS. Microglia: the missing link to decipher and therapeutically control MS progression? Int J Mol Sci. 2021;22(7):3461. doi:10.3390/ijms22073461